مجموعة فردانية
| جزء من سلسلة مقالات عن |
| علم الأنساب الوراثي |
|---|
| مفاهيم |
| موضوعات متعلقة |
النمط الفرداني haplotype هو مجموعة من الجينات في عضية يتم توارثهم معاً من والد واحد،[1][2] والمجموعة الفردانية haplogroup (haploid من باليونانية: ἁπλούς, haploûs, "طية واحدة، بسيطة" وكلمة group بالإنجليزية)، هي مجموعة أنماط فردانية متشابهة تتشارك سلف مشترك ذو طفرة تعدد أشكال النوكليوتيدات المفردة.[3][4] بشكل أكثر تحديداً، النمط الفرداني هو مزيج من البدائل على مناطق صبغية مختلفة والتي ترتبط ارتباطاً وثيقاً وتميل إلى أن تكون موروثة معاً. حيث أن المجموعة الفردانية تتألف من أنماط فردانية متشابهة، فعادة ما يمكن التنبؤ بالمجموعة الفرادية من خلال الأنماط الفرادنية. تتعلق المجموعات الفردانية بخط قرابة مفرد، والذي عادة ما يعود إلى آلاف السنين.[5] على هذا النحو، فإن عضوية المجموعة الفردانية، من قبل أي فرد، تعتمد على نسبة صغيرة نسبياً من المادة الوراثية التي يمتلكها هذا الفرد.
كل مجموعة فردانية، تنشأ، وتظل جزءاً من، مجموعة فردانية سابقة (أو مجموعة سابقة). على هذا النحو، فإن أي مجموعة متعلقة من المجموعات الفرادية يمكن صياغتها بدقة على كمرتبية متداخلة، والذي تكون كل مجموعة (مجموعة فرادنية) أيضاً مجموعة فرعية من المجموعة الفرادية الأكبر (على العكس، في النماذج ثنائية الأبوين، مثل أشجار العائلات البشرية).
تحدد المجموعات الفردانية طبيعياً عن طريق حرف أبجدي بادئ، وتتضمن التنقيحات من مجموعة أعداد وحروف إضافية، كما في (على سبيل المثال) A → A1 → A1a.
في علم الوراثة البشري، أكثر المجموعات الفرادنية التي تم دراستها هي Y-chromosome (Y-DNA) haplogroups و[[ المجموعات الفردانية البشرية لدنا المتقدرة]]، والذي يمكن استخدامهما لتحديد المعلومات السكانية الوراثية. يمر Y-DNA بمفرده على امتداد الخط الأبوي، من الأب لابنه، بينما يمر mtDNA نزولاً في الخط الأموي، من الأم للذرية من الجنسين. لا تتغير اعادات التركيب، ولا Y-DNA and mtDNA إلا عن طريق الطفرة العفوية في كل جيل بدون تمازج بين المادة الوراثية للأبوين.
تشكل المجموعات الفردانية

المتقدرة هي عضيات خلوية صغيرة تقع في سيتوپلازم خلايا حقيقيات النوى، مثل المتقدرات الموجودة في البشر. هدفها الأساسي هو توفير الطاقة للخلية. يعتقد أن المتقدرة تُخفض سلائل البكتريا المتعايشة التي تعيش حرة في السابق. ومن الدلائل التي توضح أن المتقدرات كانت تعيش حرة في وقت ما هو احتواء كل منها على دنا خلوي، يسمى دنا المتقدرة (mtDNA)، الذي تكون بنتها أكثر شبهاً بالجراثيم عن عضيات حقيقيات النوى (انظر التشوه التعايشي). الغالبية العظمى من الدنا البشري يحتوي على كروموسومات في نواة الخلية، لكن دنا المتقدرة يعد استثناءاً.
يرث الفرد السيتوپلازم والعضيات الخلوية الخاصة به عن طريق السيتوپلازم الذي الموروث حصرياً من البويضة الأمومية (خلية البويضة)؛ يمر السائل المنوي فقط على الدنا الكروموسومي، تُهضم المتقدرة كلها في البويضة. عندما تحدث طفرة في جزيء دنا المتقدرة، تمر عندئذ هذه الطفرة في الخط الأنثوي المباشر للسلالة. تنسخ الطفرات الأخطاء في تسلسل الدنا. أخطاء الاشارة يطلق عليها single nucleotide polymorphisms (SNPs).
الكروموسومات Y البشرية هي كروموسومات جنسية محددة-ذكرية؛ جميع البشر تقريباً ممن يمتلكون الكروموسوم Y سيكونون ذكور مورفولوجياً. بالرغم من أن الكروموسومات Y تقع في نواة الخلية وتقترن بالكروموسومات X إلا أن [[إعادة التركيب الجيني|يعاد تركيبها] فقط مع الكروموسوم X عند نهايات الكروموسوم Y؛ الـ95% الباقية من الكروموسوم Y لا يعاد تركيبها. هذا يعني أن الكروموسوم Y ودنا المتقدرة يتشاركان خصائص محددة.
الكروموسومات الأخرى، الأوتوسومات والكروموسوم X لدى النساء، يتشاركان المواد الوراثية الخاصة بهما (تسمى العبور الذي يؤدي إلى اعادة التركيب) أثناء الانتصاف (نوع خاص من الانقسام الخلوي الذي يحدث بغرض التكاثر الجنسي). هذا يعني بشكل قاطع أن المادة الوراثية التي تنتقل من هذه الكروموسومات تختلط في كل جيل، وهكذا يتم نقل أي طفرات جديدة بشكل عشوائي من الآباء إلى النسل.
السمة الخاصة التي تظهرها الكروموسومات Y ودنا المتقدرة تظهر في إمكانية تراكم الطفرات على جزء معين من كلا الجزيئين وأن هذه الطفرات تبقى ثابتة في مكانها على الدنا. علاوة على ذلك، فإن التسلسل التاريخي لهذه الطفران يمكن تخمينه أيضاً. على سبيل المثال، إذا احتوت مجموعة من عشرة كروموسومات Y (مأخوذة من عشرة رجال مختلفين) على طفرة، A، لكن خمسة من هذه الكروموسومات فقط تحتوي على طفرة ثانوية، B، يجب في هذه الحالة أن تكون الطفرة B قد حدثت بعد الطفرة A.
بالإضافة إلى ذلك، فن الرجال العشرة جميعهم ممن يحملون الكروموسوم ذو الطفرة A هم نسل الخط الذكوري المباشر لنفس الرجل الذي كان أول شخص يحمل هذه الطفرة. الرجل الأول الذي يحمل الطفرة B كان أيضاً نسل الخط الذكوري المباشر لهذا الرجل، لكنه أيضاً نسل للخط الذكوري المباشر لجميع الرجال الحاملين للطفرة B. سلسلة الطفرات هذه تشكل الأنساب الجزيئية. علاوة على ذلك، فإن كل طفرة تُحدد بمجموعة معينة من الكروموسومات Y تسمى المجموعة الفردانية.
جميع الرجال الحاملين للطفرة A من مجموعة فردانية مفردة، وجميع الرجال الحاملين للطفرة B هم جزء من هذه المجموعة الفردانية، لكن الطفرة B تُحدد أيضاً مجموعة فردانية إضافية (والتي تعتبر مجموعة فرعية) في مجموعتها والتي لا ينتمي إليها الرجال الحاملين للطفرة A. دنا المتقدرة والكروموسومات Y مضمنين في الأنساب والمجموعات الفردانية؛ والتي عادة ما تمثل كشجرة شبيهة بالمخططات.
علم الوراثة السكاني للمجموعات الوراثية
يفترض عادة أن هناك انتخاب طبيعي ضئيل مع أو ضد طفرة نمط فرداني معين التي ظلت باقية حتى يومنا هذا[بحاجة لمصدر]، بعيداً عن معدلات الطفرات (والتي قد تختلف من علامة لأخرى) فإن المحرك الرئيسي لعلم الوراثة السكاني المؤثر على نسب الأنماط الفردانية في مجموعة سكانية ما هو الانحراف الوراثي- التقلب العشوائي الذي يحدث بسبب عشوائية أخذ العينة التي يمرر أفرادها من المجموعة السكانية حمضهم النووي إلى أعضاء الجيل التالي من الجنس المناسب.
يؤدي هذا إلى انتشار علامة معينة ضمن مجموعة سكانية ما، حتى تصل إلى 100%، أو تختفي من السكان بالكامل. في المجموعة السكانية الكبيرة المختلطة بفعالية فإن معدل الانحراف الوراثي للألايل الشائعة يكون منخفضاً للغاية؛ ومع ذلك ، يمكن أن تتغير النسب بسرعة أكبر في مجموعة سكانية هجين صغيرة للغاية. التغيرات والتركيزات الجغرافية الملحوظة في أنماط فردانية ومجموعات معينة من الأنماط الفردانية تشهد على الآثار المميزة لعنق الزجاجة السكانية أو الأحداث المؤسسة المتكررة التي يعقبها فصل وزيادة سكانية.
السلالات التي يمكن تعبقها من الوقت الحاضر لا يمكن أن تعكس التغير الوراثي الكامل للمجموعات السكانية الأقدم: الانحراف الوراثي يعني أن بعض المتغيرات قد اختفت. تكلفة اختبارات تسلسل دنا-واي ودنا المتقدرة الكاملة حد من توافر البيانات؛ ومع ذلك، فإن تكلفتها انخفضت بشكل كبير في العقد الأخير. تحمل فترات التحام والانتشار الجغرافي الحالي للنمط الفرداني الكثير من الشكوك الخاطئة. ويعتبر هذا الأمر مزعجاً بشكل خاص في أوقات الالتحام، لأن معظم علماء الوراثة السكانية لا زالوا مستمرين (وإن كان انخفض بشكل قليل) في استخدام "طريقة ژيڤوتوڤسكي"، والتي انتقدت بشكل كبير من قبل علماء وراثة الدنا لعدم صدقها.[بحاجة لمصدر] الزنبورAngiopolybia pallens يمثل 8 مجموعات فردانية تبعاً لموقعه. يظهر هذا فكرة الانحراف الوراثي.
المجموعات الفردانية لدنا الصبغي Y
المجموعات الفردانية لدنا الصبغي Y البشري سميت من الحرف A إلى T، وتُقسم كل مجموعة منها باستخدام الأرقام والحروف الصغيرة. تسميات المجموعات الفردانية للصبغي y يتم إنشائها بواسطة اتحاد علماء الصبغي Y.[6]
| أحدث واي-سلف مشترك | |||||||||||||||||||||||||||||||
| | | |||||||||||||||||||||||||||||||
| A | BT | ||||||||||||||||||||||||||||||
| | | |||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||
| | | |||||||||||||||||||||||||||||||
| CF | DE | ||||||||||||||||||||||||||||||
| | | | | ||||||||||||||||||||||||||||||
| C | F | D | E | ||||||||||||||||||||||||||||
| | | |||||||||||||||||||||||||||||||
| G | H | IJK | |||||||||||||||||||||||||||||
| | | |||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||
| | | | | ||||||||||||||||||||||||||||||
| I | J | L | M | NO | P | S | T | ||||||||||||||||||||||||
| | | | | ||||||||||||||||||||||||||||||
| N | O | Q | R | ||||||||||||||||||||||||||||
آدم صبغي Y هو الاسم الذي أطلقه الباحثون على الذكر السلفا لأبوي الشائع الأكثر حداثة (السلالة-الذكرية) لكل البشر على قيد الحياة.
المجموعات الفردانية الرئيسية للصبغي Y، ومناطق تواجدهم الجغرافية (قبل الاستيطان الأوروبي الحديث)، تضم:
 المجموعات الفردانية المهيمن عليها الصبغي Y في التجمعات السكنية في العالم قبل العصر الاستعماري، مع دروب الهجرة المحتملة حسب نموذج الهجرة الساحلية. مجموعات بدون تحور M168
مجموعات بتحور M168(الطفرة M168 حدثت بمعدل ~50,000 bp)
مجموعات بتحور M89(الطفرة M89 حدثت بمعدل ~45,000 bp)
مجموعات بالتحورات L15 & L16
مجموعات بتحور M9(الطفرة M9 حدثة بمعدل ~40,000 bp)
مجموعات بتحور M526
|
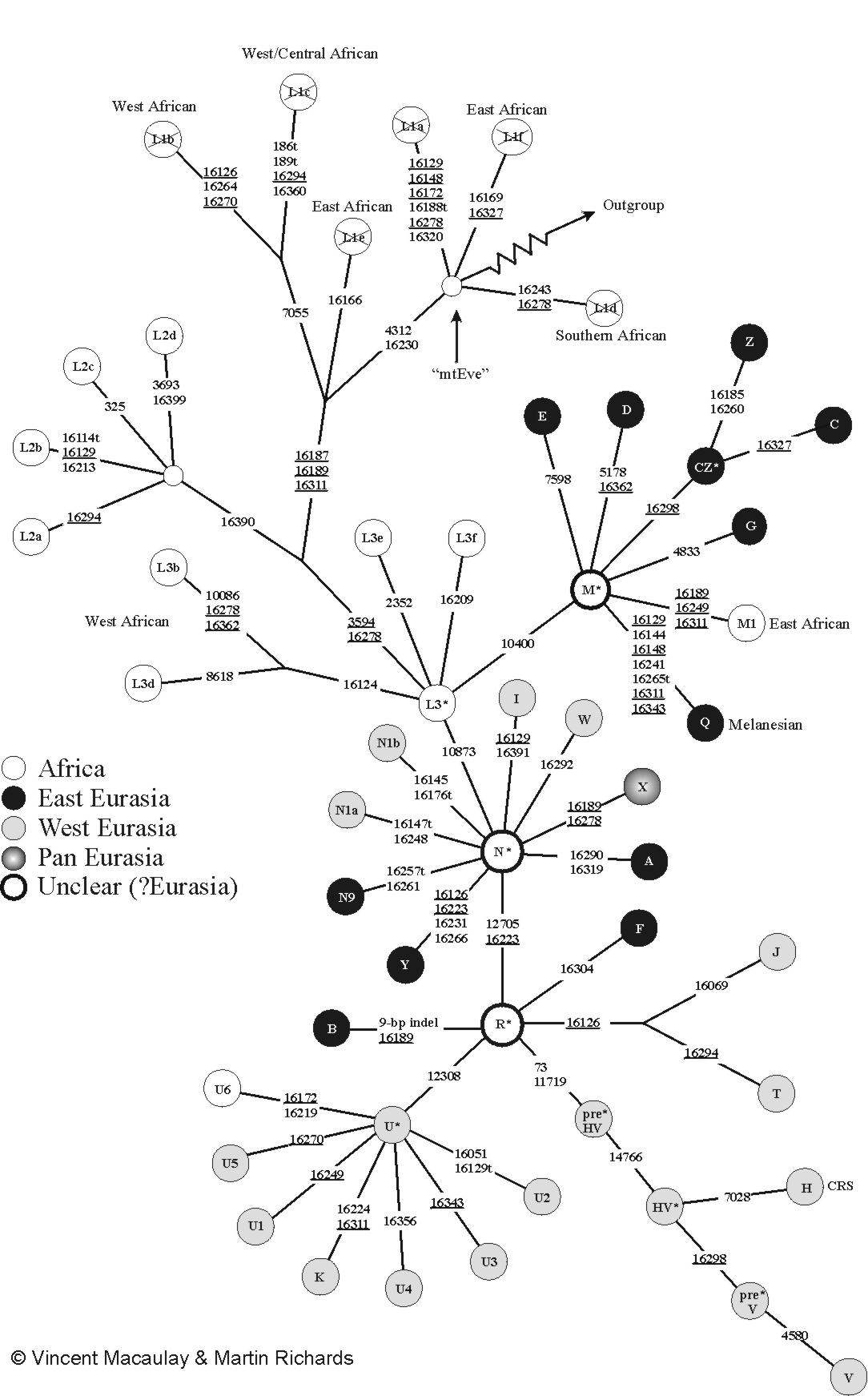
المجموعات الفردانية البشرية لدنا المتقدرة

{kind=link}
{kind=link}
{kind=link}
المجموعات الفردانية البشرية لدنا المتقدرة مرتباً أبجدياً كالتالي: A، B، C، CZ، D، E، F، G، H، HV، I، J، pre-JT، JT، K، L0، L1، L2، L3، L4، L5، L6، M، N، P، Q، R، R0، S، T، U، V، W، X، Y، و Z. أحدث نسخة من شجرة دنا المتقدرة عدلها مانيس ڤان أوڤن على موقع PhyloTree website.[8]
|
الشجرة الوراثية العرقية للمجموعات الفردانية البشرية لدنا (mtDNA) المتقدرة | |||||||||||||||||||||||||||||||||||||||
| حواء المتقدرة (L) | |||||||||||||||||||||||||||||||||||||||
| L0 | L1–6 | ||||||||||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | ||||||||||||||||||||||||||||||||||
| M | N | ||||||||||||||||||||||||||||||||||||||
| CZ | D | E | G | Q | O | A | S | R | I | W | X | Y | |||||||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | P | U | ||||||||||||||||||||||||||||||||
| HV | JT | K | |||||||||||||||||||||||||||||||||||||
| H | V | J | T | ||||||||||||||||||||||||||||||||||||
حواء المتقدرة هو الاسم الذي أطلقه الباحثون على المرأة التي السلف الأمومي الشائع الأكثر حداثة (سلالة-أنثوية) لجميع البشر الأحياء.
تعيين السكان

{kind=link}
قد تستخدم المجموعات الفردانية لتعيين السكان وراثياً وعادة ما تكون موجهة جغرافياً. على سبيل المثال، الفقرة التالية هي التقسيمات الشائعة للمجموعات الفردانية لدنا المتقدرة:
- الأفريقية: L0, L1, L2, L3, L4, L5, L6
- الغرب أوراسية: H, T, U, V, X, K, I, J, W (جميع المجموعات الفردانية الغرب أوراسية المدرجة مشتقة من المجموعة الفردانية المصغرة N)[9]
- الشرق أوراسية: A, B, C, D, E, F, G, Y, Z (ملاحظة: C, D, E, G, وZ تنتمي للمجموعة الفردانية المصغرة M)
- الأمريكية الأصلية: A, B, C, D, X
- الأسترالاسية-الملانزية: P, Q, S
تنقسم المجموعات الفردانية المتقدرة إلى مجموعات رئيسية، والتي تُعين بالحروف الثلاث المتوالية L, M, N.
الانقسام البشري الأول داخل مجموعة L بين L0 وL1-6. تسببت L1-6 في ارتفاع المجموعت L الأخرى، ومنها L3، وانقسمت إلى المجموعة M وN. تتألف المجموعة M من الموجة الأولى للهجرة البشرية خارج أفريقيا، والتي سلكت مساراً شرقياً على امتداد المناطق الساحلية الجنوبية.
يتواجد السكان المنتمون إلى المجموعة الفردانية M عبر شرق أفريقيا، آسيا، الأمريكتين، ومالنزيا. قد تمثل المجموعة N موجة هجرة أخرى خارج أفريقيا، والتي اتخذت الاتجاه الشمالي بدلاً من الشرقي. بعد فترة وجيزة من الهجرة، انقسمت مجموعة R الكبرى عن المجموعة N.
تتألف المجموعة R من مجموعتين فرعيتين تم تحديدهما على أساس توزيعها الجغرافي، توجد احداهما في جنوب شرق آسيا وأوقيانوسيا وتضم الأخرى جميع سكان أوروپا المعاصرين. المجموعة الفردانية N(xR)، أي دنا المتقدرة التي تنتمي إلى المجموعة N لكنها لا تنتمي إلى مجموعتها الفرعية R، هي مجموعة نموذجية للسكان الأصليين الأستراليين، في حين أنها موجودة أيضًا في الترددات المنخفضة بين العديد من سكان أوراسيا والأمريكتين.
المجموعة L يتألف من جميع الأفارقة تقريباً.
المجموعة M تتألف من:
M1- السكان الإثيوپيين، الصوماليين والهنود. على الأرجح بسبب المزيد من التدفق الجيني بين منطقة القرن الإفريقي وشبه الجزيرة العربية (السعودية، اليمن، عُمان)، والتي لا يفصلها سوى مضيق ضيق بين البحر الأحمر وخليج عدن.
CZ- معظم السيبريانيين؛ الفرع C- بعض الأمرينديين؛ الفرع Z- معظم السآميين، بعض الكوريين، وبعض الصينيين الشماليين، بعض سكان آسيا الوسطى.
D- بعض الأمرينديين، الكثير من سكان سيبريا وشمال شرق آسيا.
E- الملايو، البورنيون، الفلپينيون، السكان الأصليون التايوانيون، سكان پاپوا غينيا الجديدة.
G- الكثير من سكان شمال سيبريا، شمال شرق آسيا، وآسيا الوسطى.
Q- الملانزيون، الپولينزيون، سكان غينيا الجديدة.
المجموعة N تتألف من:
A- موجودة في بعض الأمرينديين، اليابانيين، والكوريين.
I- 10% من سكان شمال وشرق أوروپا.
S- بعض السكان الأصليين الأستراليين
W- بعض سكان شرق أوروپا، جنوب آسيا، وجنوب شرق آسيا.
X- الأمرينديون، سكان جنوب سيريا، جنوب غرب آسيا، وجنوب أوروپا.
Y- معظم النيڤيون والكثير من الأينوس؛ 1% في جنوب سيبريا.
R- مجموعة كبرى توجد ضمن النوع N. يمكن تقسيم السكان الموجودون فيها جغرافياً إلى غرب أوراسيا وشرق أوراسيا. معظم سكان أوروپا وعدد كبير من سكان الشرق الأوسط المعاصر يقعون ضمن هذا القسم. نسبة مئوية أصغر ضمن مجموعات النوع N. (انظر أدناه).
B- بعض الصينيين، التبتيين، المنغوليين، سكان آسيا الوسطى، الكوريين، الأمرينديين، سكان جنوب سيريا، اليابانيين، والأسترونيزيين.
F- تتواجد بصفة آساسية في جنوب شرق آسيا، وخاصة في ڤيتنام؛ 8.3% من سكان جزر هڤار في كرواتيا.[10]
R0- يتواجدون في شبه الجزيرة العربية وبين السكان الإثيوپيين والصوماليين؛ الفرع HV (الفرع H؛ الفرع V)؛ أوروپا، غرب ىسيا، شمال أفريقيا.
Pre-JT- في الشام (منطقة لبنان المعاصرة) في 25% من السكان البدو؛ الفرع JT (الفرع J؛ الفرع T)- شمال، شرق أوروپا، البحر المتوسط.
U- أعداد كبيرة في غرب أوراسيا، شبه القارة الهندية، والجزائر، ومن الهند حتى المتوسط وحتى بقية أوروپا؛ U5 تظهر بصفة خاصة التردد المرتفع في اسكنديناڤيا وبلدان البلطيق مع تردد أعلى ضمن الشعوب السامية.
التداخل بين المجموعة الفردانية-Y والمجموعة الفرادنية-mt
نطاقات التداخل بين المجموعات الفردانية-Y والمجموعات الفردانية mt المحددة، تبين السكان الذين لديهم مزيج محدد من المجموعة الفردانية-Y والمجموعة الفرادنية-mt. ليس من الضرورة أن تحدث الطفرات Y والطفرات mt في الوقت نفسه، والمعدلات التفاضلية للانتقاء الجنسي بين الجنسين تندمج مع تأثير المؤسس والانحراف المراثي الذي يمكن أن يغير تركيب المجموعة الفردانية للسكان، ومن ثم يكون التدخل حاداً.
التداخلات شديدة الحدة بين المجموعات الفردانية Y-DNA والمجموعات الفردانية mtDNA كالتالي:
| المجموعة الفردانية Y-DNA | المجموعة الفردانية mtDNA | المنطقة الجغرافية و/أم الشعوب |
|---|---|---|
| A | L0 | شرق وجنوب أفريقيا |
| B | L1, L4 | شرق ووسط أفريقيا |
| E | L2, L3 | جميع أنحاء القارة الأفريقية |
| D, O, N, C2 (كان يعرف باسم C3) | CZ/C/Z, D, G (الأنواع M); A, N9/Y (الأنواع N)؛ B, F (الأنواع R) | شرق آسيا، سيبريا |
| K2b1, C1 (كان يعرف في السابق باسم CxC3), PxQR (في تيمور ونيگريتوس من الفلپين) | B, P (الأنواع R)؛ N؛ Q (النوع M) بالإضافة لمجموعات فرعية أوقيانوسية محددة من النوع M | أوقيانوسيا |
| R, I, T, J, E (V13, M81 والأنواع M123) | R0, HV/H/V, JT/J/T, U/K (الأنواع R types), M1 (M) | أوروپا، غرب آسيا، شمال أفريقيا |
| H, R1a-z93, R2, L | U2, U7 أنواع فرعية أخرى من R، أنواع فرعية من M. | جنوب آسيا |
| Q, C2 (كانت تعرف باسم C3) | A, X (الأنواع N); C, D (الأنواع M) | أقصى شرق سيبيريا، الأمريكتان |
تعيين الكروموسوم-Y والمجموعة الفردانية الجغرافية MtDNA
الفقرة التالية هي قائمة لتعيين الكروموسوم-Y والمجموعة الفردانية الجغرافية MtDNA والمقترحة من قبل بكادا وزملائه، 2013.[11]
الكروموسوم-Y
تبعاً للمجموعات الفردانية SNPS والتي تميل إلى تحديد عمر حدث الانقراض الأول حوالي 45-50 مليون سنة مضت. يبدو أن المجموعات الفردانية في حدث الانقراض الثاني قد انقسمت منذ 32-35 مليون سنة مضت حسب حضارة Mal'ta. يظهر حدث الانقراض الصفري البري to be Toba والذي يتضح أن المجموعة الفردانية CDEF* قد انقسمت أثناؤه إلى المجموعات C، DE وF. لا يوجد بين المجموعة C وF أي قاسم مشترك بينما المجموعة D وE لديها الكثير من القواسم المشتركة. حدث الانقراض الأول تبعاً للتقديرات الحالية وقع بعد Toba، بالرغم من أن الدنا العتيق الأقدم يمكن أن يدفع بحدث الانقراض الصفري البري إلى فترة أبعد من Toba، ويدفع حدث الانقراض الأول هنا إلى ما وراء Toba. المجموعات الفردانية ذات أحداث الانقراض والتي تم رصدها من قبلهم لديها أصل مشكوك فيه، وهذا لأن أحداث الانقراض تؤدي إلى اختناقات شديدة، جميع الملاحظات التي تقدمها هذه المجموعات هي مجرد تخمينات. لاحظ أن استمرار المجموعة SNP من الدنا العتيق يمكن أن يختلف بشكل كبير مما يعني أنه على الرغم من انقسام جميع هذه المجموعات في الوقت نفسه فلا أحد يعرف متى حدث هذا الانقسام.[12][13]
| الأصل | المجموعة الفردانية | الدلالة |
| أوروپا | E1b1b1a2 | V13 |
| أوروپا (حدث الانقراض الثاني؟) | I | M170, M253, P259, M227, M507 |
| أوروپا | I1b | P215,M438, P37.2, M359, P41.2 |
| أوروپا | I1b2 | M26 |
| أوروپا | I1c | M223, M284, P78, P95 |
| أوروپا | J2a1 | M47 |
| أوروپا | J2a2 | M67, M166 |
| أوروپا | J2a2a | M92 |
| أوروپا | J2b | M12,M102, M280, M241 |
| أوروپا | R1b1b1a | M412, P310 |
| أوروپا | R1b1b1a1 | L11 |
| أوروپا | R1b1b1a1a | U106 |
| أوروپا | R1b1b1a1b | U198, P312, S116 |
| أوروپا | R1b1b1a1b1 | U152 |
| أوروپا | R1b1b1a1b2 | M529 |
| أوروبا | R1b1b1a1b3,4 | M65, M153 |
| أوروپا | R1b1b1a1b5 | SRY2627 |
| جنوب آسيا أو ملانسيا | C1(formerly known as CxC3) | Z1426 |
| شمال آسيا | C2 (كانت تعرف C3) | M217+ |
| إندونسيا أو جنوب آسيا (حدث الانقراض الأول؟) | F | M89, M282 |
| أوروپا (القوقاز) (حدث الانقراض الثاني؟) | G | M201, M285, P15, P16, M406 |
| جنوب آسيا | H | M69, M52, M82, M197, M370 |
| أوروپا أو الشرق الأوسط (حدث الانقراض الثاني؟) | J1 | M304, M267, P58, M365, M368, M369 |
| أوروپا أو الشرق الأوسط (حدث الانقراض الثاني؟) | J2 | M172, M410, M158, M319, DYS445=6, M339, M340 |
| غرب بورما في أوراسيا (حدث الانقراض الأول؟)[14] | ||
| إندونسيا (حدث الانقراض الأول؟) [14] | K2 (NOPS) | M526 |
| جنوب آسيا | L | M11, M20, M27, M76, M317, M274, M349, M357 |
| شرق آسيا، جنوب شرق آسيا | N | M231, M214, LLY22g, Tat, M178 |
| شرق آسيا، جنوب شرق آسيا، جنوب آسيا (حدث الانقراض الثاني؟) | O | M175, M119 |
| إندونسيا، الفلپين (حدث الانقراض الأول؟) | P (xQR) | 92R7, M207, M173, M45 |
| جنوب آسيا، سيبريا (حدث الانقراض الثاني؟) | R وQ (QR) انفصلا [14] | MEH2, M242, P36.2, M25, M346 |
| الشرق الأوسط، أوروپا، سبيريا، جنوب آسيا | R1a1 | M420, M17, M198, M204, M458 |
| الأناضول، جنوب شرق أوروپا ؟ | R1b | M173, M343, P25, M73 |
| أوروپا | R1b1b | M269 |
| أوروپا | R1b1b1 | L23 |
| پاكستان، الهند (حدث الانقراض الثاني؟) | R2 | M479, M124 |
| الشرق الأوسط | T | M70 |
| شمال أفريقيا | E1b1b1a | M78 |
| شمال أفريقيا | E1b1b1a1 | V12 |
| شمال أفريقيا | E1b1b1a1b | V32 |
| شمال أفريقيا | E1b1b1a3 | V22 |
| شمال أفريقيا | E1b1b1a4 | V65 |
| شمال أفريقيا | E1b1b1b | M81 |
| غرب أفريقيا، شمال أفريقيا | A | M91, M13 |
| شرق أفريقيا | B | M60, M181, SRY10831.1, M150, M109, M112 |
| آسيا، أفريقيا | DE | M1, YAP, M174, M40, M96, M75, M98 |
| شرق آسيا | D | M174 |
| شرق أفريقيا (انفصال السلف عن E1b1a ليكون حدث الانقراض الثاني) | E1b1b1 | M35 |
| شرق أفريقيا | E1b1b1c | M123, M34 |
| غرب أفريقيا (حدث الانقراض الأول؟) | E1a | M33 |
| شرق أفريقيا (حدث الانقراض الأول هو الفصل بين E1b1 وE1a، حدث الانقراض الثاني هو الفصل بين E1b1b وE1b1a) | E1b1 | P2, M2, U175, M191 |
| الشرق الأوسط | R1b1a | V88, M18 |
mtDNA
| الأصل | المجموعة الفردانية |
| أوروپا | H1 |
| أوروپا | H11a |
| أوروپا | H1a |
| أوروپا | H1b |
| أوروپا | H2a |
| أوروپا | H3 |
| أوروپا | H5a |
| أوروپا | H6a |
| أوروپا | H7 |
| أوروپا | HV0/HV0a/V |
| أوروپا | I4 |
| أوروپا | J1c7 |
| أوروپا | J2b1 |
| أوروپا | T2b* |
| أوروپا | T2b4 |
| أوروپا | T2e |
| أوروپا | U4c1 |
| أوروپا | U5* |
| أوروپا | U5a |
| أوروپا | U5a1b1 |
| أوروپا | U5b* |
| أوروپا | U5b1b* |
| أوروپا | U5b1c |
| أوروپا | U5b3 |
| أوروپا | X2c'e |
| الشرق الأوسط | I |
| الشرق الأوسط | A |
| الشرق الأوسط | B |
| الشرق الأوسط | C/Z |
| الشرق الأوسط | D/G/M9/E |
| الشرق الأوسط | F |
| الشرق الأوسط | H* |
| الشرق الأوسط | H13a1 |
| الشرق الأوسط | H14a |
| الشرق الأوسط | H20 |
| الشرق الأوسط | H2a1 |
| الشرق الأوسط | H4 |
| الشرق الأوسط | H6b |
| الشرق الأوسط | H8 |
| الشرق الأوسط | HV1 |
| الشرق الأوسط | I1 |
| الشرق الأوسط | J / J1c / J2 |
| الشرق الأوسط | J1a'b'e |
| الشرق الأوسط | J1b1a1 |
| الشرق الأوسط | J1b2a |
| الشرق الأوسط | J1d / J2b |
| الشرق الأوسط | J1d1 |
| الشرق الأوسط | J2a |
| الشرق الأوسط | J2a2a1 |
| الشرق الأوسط | K* |
| الشرق الأوسط | K1a* |
| الشرق الأوسط | K1b1* |
| الشرق الأوسط | N1a* |
| الشرق الأوسط | N1b |
| الشرق الأوسط | N1c |
| الشرق الأوسط | N2 |
| الشرق الأوسط | N9 |
| الشرق الأوسط | R* |
| الشرق الأوسط | R0a |
| الشرق الأوسط | T |
| الشرق الأوسط | T1* |
| الشرق الأوسط | T1a |
| الشرق الأوسط | T2 |
| الشرق الأوسط | T2c |
| الشرق الأوسط | T2i |
| الشرق الأوسط | U1* |
| الشرق الأوسط | U2* |
| الشرق الأوسط | U2e |
| الشرق الأوسط | U3* |
| الشرق الأوسط | U4 |
| الشرق الأوسط | U4a* |
| الشرق الأوسط | U7 |
| الشرق الأوسط | U8* |
| الشرق الأوسط | U9a |
| الشرق الأوسط | X |
| الشرق الأوسط | X1a |
| الشرق الأوسط | X2b1 |
| شمال أفريقيا | L3e5 |
| شمال أفريقيا | M1 |
| شمال أفريقيا | M1a1 |
| شمال أفريقيا | U6a |
| شمال أفريقيا | U6a1'2'3 |
| شمال أفريقيا | U6b'c'd |
| شرق أفريقيا | L0* |
| شرق أفريقيا | L0a1 |
| شرق أفريقيا | L0a1b |
| شرق أفريقيا | L0a2* |
| شرق أفريقيا | L3c/L4/M |
| شرق أفريقيا | L3d1a1 |
| شرق أفريقيا | L3d1d |
| شرق أفريقيا | L3e1* |
| شرق أفريقيا | L3f* |
| شرق أفريقيا | L3h1b* |
| شرق أفريقيا | L3i* |
| شرق أفريقيا | L3x* |
| شرق أفريقيا | L4a'b* |
| شرق أفريقيا | L5* |
| شرق أفريقيا | L6 |
| شرق أفريقيا | N* / M* / L3* |
| غرب أفريقيا | L1b* |
| غرب أفريقيا | L1b3 |
| غرب أفريقيا | L1c* |
| غرب أفريقيا | L1c2 |
| غرب أفريقيا | L2* |
| غرب أفريقيا | L2a |
| غرب أفريقيا | L2a1* |
| غرب أفريقيا | L2a1a2'3'4 |
| غرب أفريقيا | L2a1b |
| غرب أفريقيا | L2a1b'f |
| غرب أفريقيا | L2a1c1'2 |
| غرب أفريقيا | L2a1(16189) |
| غرب أفريقيا | L2a2 |
| غرب أفريقيا | L2b* |
| غرب أفريقيا | L2c1'2 |
| غرب أفريقيا | L2d |
| غرب أفريقيا | L2e |
| غرب أفريقيا | L3b |
| غرب أفريقيا | L3b1a3 |
| غرب أفريقيا | L3b(16124!) |
| غرب أفريقيا | L3b2a |
| غرب أفريقيا | L3d* |
| غرب أفريقيا | L3e2'3'4 |
| غرب أفريقيا | L3f1b* |
انظر أيضاً
- المشروع الدولي لخريطة المجموعات الفردانية
- تطور جزيئي
- نسالة جزئية
- نمط فرداني Haplotype
- علم الوراثة التطوري البشري
- عرق (علم الأحياء) / عرق (تصنيف بشري)
- علم الأنساب الوراثي
- تحليل الجينات
- موروثة
- قائمة موضوعات علم الأنساب الوراثي
- قائمة المجموعات الفدرانية للمشاهير
- مجموعة فرعية
مراجع
- ^ By C. Barry Cox, Peter D. Moore, Richard Ladle. Wiley-Blackwell, 2016. ISBN 978-1-118-96858-1 p106. Biogeography: An Ecological and Evolutionary Approach
- ^ Editorial Board, V&S Publishers, 2012, ISBN 9381588643 p137.Concise Dictionary of Science
- ^ Arora, Devender; Singh, Ajeet; Sharma, Vikrant; Bhaduria, Harvendra Singh; Patel, Ram Bahadur (2015). "Hgs Db: Haplogroups Database to understand migration and molecular risk assessment". Bioinformation. 11 (6): 272–5. doi:10.6026/97320630011272. PMC 4512000. PMID 26229286.
- ^ International Society of Genetic Genealogy 2015 Genetics Glossary
- ^ "Haplogroup definition in DNA--NEWBIE GLOSSARY". The International Society of Genetic Genealogy.
- ^ "Y Chromosome Consortium".
- ^ Rootsi S, Magri C, Kivisild T, Benuzzi G, Help H, Bermisheva M, Kutuev I, Barać L, Pericić M, Balanovsky O, Pshenichnov A, Dion D, Grobei M, Zhivotovsky LA, Battaglia V, Achilli A, Al-Zahery N, Parik J, King R, Cinnioğlu C, Khusnutdinova E, Rudan P, Balanovska E, Scheffrahn W, Simonescu M, Brehm A, Goncalves R, Rosa A, Moisan JP, Chaventre A, Ferak V, Füredi S, Oefner PJ, Shen P, Beckman L, Mikerezi I, Terzić R, Primorac D, Cambon-Thomsen A, Krumina A, Torroni A, Underhill PA, Santachiara-Benerecetti AS, Villems R, Semino O (Jul 2004). "Phylogeography of Y-chromosome haplogroup I reveals distinct domains of prehistoric gene flow in europe" (PDF). American Journal of Human Genetics. 75 (1): 128–37. doi:10.1086/422196. PMC 1181996. PMID 15162323.
- ^ "PhyloTree.org".
- ^ Loogväli EL, Roostalu U, Malyarchuk BA, Derenko MV, Kivisild T, Metspalu E, et al. (2004). "Disuniting uniformity: a pied cladistic canvas of mtDNA haplogroup H in Eurasia". Mol. Biol. Evol. 21 (11): 2012–21. doi:10.1093/molbev/msh209. PMID 15254257.
- ^ Tolk HV, Barac L, Pericic M, Klaric IM, Janicijevic B, Campbell H, Rudan I, Kivisild T, Villems R, Rudan P (Sep 2001). "The evidence of mtDNA haplogroup F in a European population and its ethnohistoric implications". European Journal of Human Genetics. 9 (9): 717–23. doi:10.1038/sj.ejhg.5200709. PMID 11571562.
- ^ Bekada A, Fregel R, Cabrera VM, Larruga JM, Pestano J, Benhamamouch S, González AM (2013). "Introducing the Algerian mitochondrial DNA and Y-chromosome profiles into the North African landscape". PLOS ONE. 8 (2): e56775. doi:10.1371/journal.pone.0056775. PMC 3576335. PMID 23431392.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ "Common genetic ancestors lived during roughly same time period". 1 Aug 2013. Retrieved 23 Jan 2015.
- ^ Raghavan M, Skoglund P, Graf KE, Metspalu M, Albrechtsen A, Moltke I, Rasmussen S, Stafford TW, Orlando L, Metspalu E, Karmin M, Tambets K, Rootsi S, Mägi R, Campos PF, Balanovska E, Balanovsky O, Khusnutdinova E, Litvinov S, Osipova LP, Fedorova SA, Voevoda MI, DeGiorgio M, Sicheritz-Ponten T, Brunak S, Demeshchenko S, Kivisild T, Villems R, Nielsen R, Jakobsson M, Willerslev E (Jan 2014). "Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans". Nature. 505 (7481): 87–91. doi:10.1038/nature12736. PMC 4105016. PMID 24256729.
- ^ أ ب ت Karafet TM, Mendez FL, Sudoyo H, Lansing JS, Hammer MF (Mar 2015). "Improved phylogenetic resolution and rapid diversification of Y-chromosome haplogroup K-M526 in Southeast Asia". European Journal of Human Genetics. 23 (3): 369–73. doi:10.1038/ejhg.2014.106. PMC 4326703. PMID 24896152.
وصلات خارجية
عامة
أخبار
- Indian maternal gene pool, Journal of Human Genetics
- Dienekes' Anthropology Blog frequent highlights of new results.
جميع المجموعات الفردانية للدنا
Y-Chromosome - *http://www.scs.uiuc.edu/~mcdonald/WorldHaplogroupsMaps.pdf
المجموعات الفردانية لدنا الكروموسوم Y
- Y Chromosome Consortium
- ISOGG Y-DNA Haplogroup Tree
- PhyloTree's Y-tree A minimal reference phylogeny for the human Y-chromosome
- Haplogroup Predictor
- The Y Chromosome Consortium (2002), A Nomenclature System for the Tree of Human Y-Chromosomal Binary Haplogroups, Genome Research, Vol. 12(2), 339-348, February 2002. (Detailed hierarchical chart has conversions from previous naming schemes)
- Semino et al. (2000), The Genetic Legacy of Paleolithic Homo sapiens sapiens in Extant Europeans, Science, Vol 290 (paper which introduced the "Eu" haplogroups).
- Y-DNA Ethnographic and Genographic Atlas and Open-Source Data Compilation
المجموعات الفردانية لدنا المتقدرة
- PhyloTree - The phylogenetic tree of global human mitochondrial DNA variation[1]
- PhyloD3 - D3.js-based phylogenetic tree based on PhyloTree
- MitoTool - a web server for the analysis and retrieval of human mitochondrial DNA sequence variations
- HaploGrep - automatic classification of mitochondrial DNA haplogroups based on PhyloTree
- HaploFind - fast automatic haplogroup assignment pipeline for human mitochondrial DNA
- graphical mtDNA haplogroup skeleton
- The Making of the African mtDNA Landscape
- Do the Four Clades of the mtDNA Haplogroup L2 Evolve at Different Rates?
{kind=link}
برامج حاسوبية
المصادر
|
الشجرة الوراثية العرقية للمجموعات الفردانية البشرية لدنا (mtDNA) المتقدرة | |||||||||||||||||||||||||||||||||||||||
| حواء المتقدرة (L) | |||||||||||||||||||||||||||||||||||||||
| L0 | L1–6 | ||||||||||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | ||||||||||||||||||||||||||||||||||
| M | N | ||||||||||||||||||||||||||||||||||||||
| CZ | D | E | G | Q | O | A | S | R | I | W | X | Y | |||||||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | P | U | ||||||||||||||||||||||||||||||||
| HV | JT | K | |||||||||||||||||||||||||||||||||||||
| H | V | J | T | ||||||||||||||||||||||||||||||||||||
| أحدث واي-سلف مشترك | |||||||||||||||||||||||||||||||
| | | |||||||||||||||||||||||||||||||
| A | BT | ||||||||||||||||||||||||||||||
| | | |||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||
| | | |||||||||||||||||||||||||||||||
| CF | DE | ||||||||||||||||||||||||||||||
| | | | | ||||||||||||||||||||||||||||||
| C | F | D | E | ||||||||||||||||||||||||||||
| | | |||||||||||||||||||||||||||||||
| G | H | IJK | |||||||||||||||||||||||||||||
| | | |||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||
| | | | | ||||||||||||||||||||||||||||||
| I | J | L | M | NO | P | S | T | ||||||||||||||||||||||||
| | | | | ||||||||||||||||||||||||||||||
| N | O | Q | R | ||||||||||||||||||||||||||||
- All articles lacking reliable references
- Articles lacking reliable references from September 2016
- CS1 maint: unflagged free DOI
- Articles containing Greek-language text
- Articles with unsourced statements from March 2012
- Articles with unsourced statements from July 2011
- دنا
- تطور بشري
- علم الوراثة العرقي
- علم الوراثة السكاني
- علم الوراثة الكلاسيكي